
¿Por qué el cerebro controla mitades opuestas de nuestro cuerpo?

Cada mitad de nuestro cerebro es responsable de mitades opuestas de nuestro cuerpo (hemicuerpos). Esto ocurre porque gran parte de las fibras nerviosas dirigidas al resto del cuerpo decusan, es decir, cambian de lado. Aunque esto refleja el tipo de conexiones que tenemos, no explica por qué.
Frecuencia
Esto no es exclusivo de los humanos, pero tampoco es un fenómeno tan común para estar presente en todos los animales con simetría bilateral (Bilateria). Es una característica única compartida por los cordados. Dado que, muy remotamente, provenimos de los invertebrados, se sugiere que en algún protocordado, hace unos 600 millones de años, se produjo el cambio. Esto genera más preguntas, porque ¿qué ventaja le podría ofrecer? Es más, si aporta un beneficio y se ha conservado en las 81000 especies del filo de los cordados, ¿por qué no se ha experimentado en millón y medio de especies repartidas en los otros 38 filos?
Descubriendo la decusación

Tracto corticoespinal
_(14579816217).jpg)
El tracto corticoespinal, principal vía del movimiento voluntario, se conocería con más detalle en los siguientes siglos. La conexión llega desde la quinta capa de neuronas de la corteza frontal y parietal, atravesando la mitad anterior del miembro posterior de la cápsula interna y, antes de llegar al tronco cerebral, forma pedúnculos cerebrales. En el bulbo raquídeo, unos milímetros por debajo del cuarto ventrículo cerebral, la mayor parte de las fibras cruzan la línea media para formar el tracto corticoespinal lateral, mientras el 10-25% (1) que continúan en el mismo sentido forman el tracto corticoespinal anterior o ventral (2). Cada uno tiene distintas longitudes. Mientra el primero llega a los segmentos sacros, con conexiones a través de motoneuronas de la parte dorsolateral del cuerno anterior (3), para controlar la motricidad fina de los miembros inferiores; el segundo se queda, en seres humanos, en segmentos torácicos superiores, ramificándose para conectar con interneuronas de ambos cuernos anteriores ventromediales, controlando la musculatura axial.
Varios cruces

Aunque la decusación se descubrió por primera vez en el tracto corticoespinal, este tan solo está presente en mamíferos y desarrollado en primates y animales que necesitan realizar movimientos precisos en las extremidades. Este desarrollo se manifiesta en más fibras, más grandes, con mayor velocidad de conducción, debido a la mielinización (4), y con conexión más directa con las motoneuronas, en lugar de conexiones multisinápticas. La presencia en estas especies muestra que este tracto es relativamente reciente. En tractos primitivos, las neuronas, en lugar de recorrer grandes distancias con su axones y requerir únicamente una o dos sinapsis o conexiones, implican a más neuronas intermedias, como en una carrera de relevos con muchos corredores más lentos donde muchos llegan al mismo sitio, pues tienen sinapsis recurrentes. Esta diferencia se observa en las crías de animales. En humanos, nuestro tracto corticoespinal se manifiesta de forma tardía en el desarrollo embrionario y continúa en nuestros primeros años, por eso, no nos lanzamos a levantarnos y correr al poco de nacer.
Los tractos reticuloespinal , vestibuloespinal y rubroespinal originados en el tronco encefálico son vías motoras más primitivas, especialmente los dos primeros, presentes en embriones de lampreas, y, estos sí, tienen más similitudes entre vertebrados. En los humanos, los tractos reticuloespinales y vestibuloespinales controlan el tono muscular, la postura y el equilibrio. Sus proyecciones neuronales son principalmente del mismo lado, actuando con las motoneuronas a través de interneuronas, pudiendo controlar miembros en algunas especies. En cambio, el tracto rubroespinal, que solo existe en especies con miembros o pseudomiembros, ocupa una posición intermedia entre estos y el corticoespinal. Por ello, como este último, la mayoría de las fibras decusan y actúan directamente sobre motoneuronas. En mamíferos, conforme más desarrollado está el tracto corticoespinal, menos presente está el rubroespinal, más importante en cuadrúpedos.
En el cerebro, las comisuras son lugares de conexión de áreas homólogas de ambos hemisferios. En mamíferos placentarios, se le suma el cuerpo calloso.¿Por qué decusar?
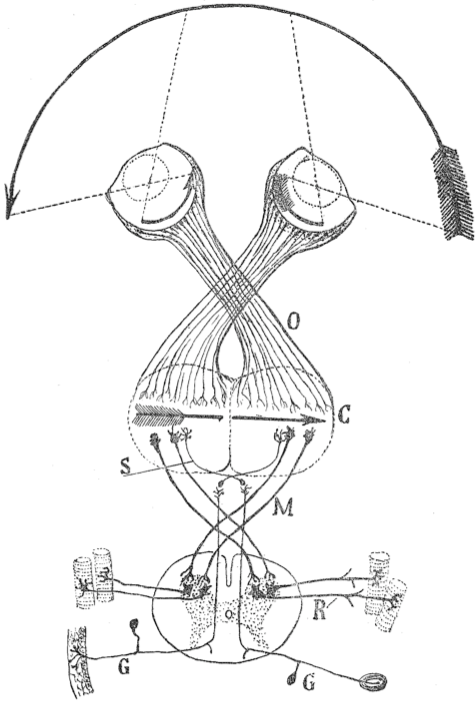
Como hemos visto, el cruce de fibras no se limita al tracto corticoespinal. Sin embargo, encontrar distintos momentos en los que ocurrió no ofrece por sí mismo una explicación. En 1899, Santiago Ramón y Cajal argumentó que el quiasma óptico, donde se cruzan las fibras de los nervios ópticos, servía para reinvertir la imagen formada sobre la retina. En especies con visión binocular, este cruce es parcial (5), permitiendo la visión estereoscópica y mejorando las habilidades motoras.
Alternativas
Aunque 25 años después Ramón y Cajal pidió mayor investigación a su teoría de la decusación compensatoria de la integración sensorial y motora, se ha mantenido vigente durante más de un siglo y sigue siendo la más aceptada. No obstante, hay detalles que no se corresponden con la realidad. Al contrario de lo que afirmó, el número de fibras que cruzan el quiasma óptico y las que decusan el tracto corticoespinal no son inversamente proporcionales.
Se han propuesto hipótesis de distinto calado para justificar este hecho. Una explicación sencilla es que proporcionan estabilidad física o que es el resultado del desarrollo de un cerebro esférico a uno simétrico y bilateral, pero las decusaciones en invertebrados son más excepciones que la norma.
Por otra parte, se alega que si un estímulo visual llega por un lado, la
información se dirigirá y se procesará en el lado opuesto del cerebro. La
reacción también sucederá en este lado opuesto al estímulo, permitiendo
contraer rápidamente la mitad del cuerpo y alejar la cabeza. Además,
cuando hay miembros, los ipsilaterales al estímulo se extenderían a través de
los tractos rubroespinales y corticoespinales. Los estímulos que provocarían
esta respuesta serían visuales y táctiles, cuyas vías se cruzarían y
derivarían en una respuesta común. La dependencia de la vista y el tacto
habría surgido en consonancia con el desarrollo de estas vías y la
psicomotricidad fina.
Asimismo, se ha propuesto la existencia potencial de vertebrados ancestrales con un solo ojo central, cuya retina fue haciéndose más cóncava, invirtiéndose la imagen proyectada y obligando a la reorganización del cerebro, llevando posteriormente a producir el quiasma óptico.
Hipótesis retorcidas
La alternativa más aceptada son las hipótesis de la torsión axial o modelo de torsión somática. Proponen que la decusación, más que una ventaja, fue una consecuencia de una reorganización corporal. En invertebrados, el neuroeje, es decir, la dirección del sistema nervioso central, comienza en una masa gangliónica o cerebro en un extremo del cuerpo, sobre la orofaringe. Sin embargo, salvo este extremo, el resto del neuroeje se traslada de su posición dorsal a una ventral. En cambio, en vertebrados, el neuroeje se mantiene dorsal. Esta diferencia con los invertebrados se observa en la notocorda, precursora de la columna vertebral, en protocordados como los tunicados, enteropneustos y amphioxus.

Aunque no lo hizo pensando en la decusación, en 1822, Étienne Geoffroy Saint-Hilaire observó al voltear cangrejos de río que, de esta manera, el neuroeje se situaba sobre el aparato digestivo y el corazón, como en los vertebrados. Incluso décadas después con la teoría de la evolución de Darwin, parecía un cambio muy brusco para el paso de invertebrados a vertebrados. No obstante, fue recordado siglo y medio después cuando se aceptó que la evolución también incluía a cambios rápidos. Esta inversión también se estudió en otras áreas, como las regiones principalmente sensitivas dorsales y motoras ventrales en cordados, cuya posición es opuesta en invertebrados, reflejándose en el amphioxus. Este fenómeno también se observó en la posición del corazón respecto al sistema circulatorio.

Por lo tanto, las hipótesis de la torsión axial postulan que el cuerpo giró sobre su eje: 180º salvo en la zona anterior a la faringe, incluyendo los órganos internos, o en dos giros opuestos de 90º en la gástrula, primero en sentido horario en la zona del prosencéfalo-ojo antes de la neurulación y posteriormente en sentido antihorario en la zona del mesencéfalo pero manteniendo el corazón y otras vísceras. En ambos modelos, la torsión habría comenzado a un nivel posterior al nervio olfatorio, primer par craneal y el más anterior, pues en el segundo (nervio óptico), cuarto (troclear), séptimo (facial) y duodécimo (hipogloso) par las fibras ya cambian de lado para llegar a sus núcleos en el lado opuesto, aunque técnicamente solo el cuarto decusa, haciéndolo en el velo medular superior. Igualmente, explicaría que cada hemisferio cerebelar represente al hemicuerpo ipsilateral.

La dificultad que encuentra esta hipótesis es explicar la decusación parcial de las fibras y que no todas las neuronas lo hacen en el quiasma óptico. No solo lo hacen también en el cuerpo calloso y las comisuras anterior y posterior, sino también las células de Müller y Mauthner del rombencéfalo, aunque lo hagan en el mismo segmento donde se originan.
Notas
- El número de fibras que decusan varían por persona e, independientemente de la dominancia, las que cruzan de izquierda a derecha son más numerosas. En este lado, las fibras llegan más lejos.
- En el cuerpo calloso decusan algunas fibras y algunas vuelven al lado ipsilateral en la médula espinal.
- Si cortamos transversalmente, en rodajas, la médula espinal, la sustancia gris central tiene una forma de mariposa o de H. Los cuernos son las puntas de las "alas" de la mariposa o los extremos de la H. Anterior o ventral es delante; posterior o dorsal es detrás; medial es próximo a la línea media y lateral es alejado a esta.
- La mielina es una sustancia aislante de los axones, la extensión larga de las neuronas, que actúa como los condensadores eléctricos y permite acelerar el impulso sin necesitar una fibra más gruesa. Por supuesto, se pueden tener fibras gruesas y mielinizadas, obteniendo los beneficios de ambos.
-
Las fibras que recogen información de la mitad lateral del ojo se dirigen a
la corteza visual del mismo lado. Las fibras que recogen información de la
mitad medial del ojo, se dirigen a la corteza visual del hemisferio opuesto.
Fuentes
- Kinsbourne, M. (2013). Somatic twist: A model for the evolution of decussation. Neuropsychology, 27(5), 511.
- Vulliemoz, S., Raineteau, O., & Jabaudon, D. (2005). Reaching beyond the midline: why are human brains cross wired?. The Lancet Neurology, 4(2), 87-99.
- Mora, C., Velásquez, C., & Martino, J. (2019). The neural pathway midline crossing theory: a historical analysis of Santiago Rámon y Cajal’s contribution on cerebral localization and on contralateral forebrain organization. Neurosurgical Focus, 47(3), E10.
- Victor, J. D., Apkarian, P., Hirsch, J., Conte, M. M., Packard, M., Relkin, N. R., ... & Shapley, R. M. (2000). Visual function and brain organization in non-decussating retinal–fugal fibre syndrome. Cerebral Cortex, 10(1), 2-22.
-
de Lussanet, M. H., & Osse, J. W. (2015). Decussation as an axial twist:
A comment on Kinsbourne (2013).





Hola, yo lo he pensado alguna vez. Creo que es un mecanismo de seguridad, como la infertilidad del cruce entre especies. Si el sistema nervioso no se cruzara en su recorrido, en pocos millones de años las especies tendrían sistemas dobles e independientes.